

۱- مقدمه
به تازگی، تقلب و دستکاری مواد غذایی به دلیل انگیزه سودجویی سریع اقتصادی به طور قابل توجهی افزایش یافته است. آنالیز مواد غذایی برای تضمین اصالت مواد غذایی ضروری است. نتایج این آنالیزها به مردم کمک میکند تا از مصرف مواد غذایی حاوی ترکیبات خاص مضر برای سلامتی مثل آلرژن ها خودداری کرده و نگرانیهای مذهبی و فرهنگی نداشته باشند. همچنین این آنالیزها به محدود کردن روشهای تقلب مانند رسوایی گوشت اسب در اروپا کمک میکند.
به عنوان مثال، در پاسخ به این رسوایی، مقامات ایمنی مواد غذایی (FSA) و وزارت محیط زیست، غذا و امور روستایی (Defra) آستانه عدم اعلام گونههای گوشت در محصولات گوشتی را در اروپا به 1٪ اعلام کردند و اکنون ضروری است که روشهای تجزیهای به اندازه کافی حساس و دقیق باشند تا بتوانند حضور مقادیر کمی از یک ماده در محصولات غذایی بررسی کنند. به طور سنتی، از آزمایشهای ایمونوسوربنت مرتبط به آنزیم (ELISA) یا آزمایشهای مبتنی بر کشت برای تعیین نوع گونه در مواد غذایی یا تشخیص آلودگی میکروبی استفاده میشود. ELISA در حالی که آسان و ارزان است، وابسته به آنتیبادیهای هدف برای بافتها/گونههای هدف است و اختصاصی بودن (گزینش پذیری) ضعیفی دارد زیرا نمی تواند بین گونه های مرتبط فیلوژنتیکی در نمونه های غذایی تمایز قائل شود.
فیلوژنی (Phylogeny) یا تبارزایش بررسی روابط بین گروههای مختلف ارگانیسمها و رشد تکاملی آنها است. این علم مبتنی بر فرضیه فیلوژنتیک است که بر طبق آن همه موجودات زنده یک جد مشترک دارند. این روابط از طریق مقایسه شباهتهای ژنتیکی و آناتومیکی موجودات زنده با یکدیگر نشان داده میشود.
"نسخه اصلی این مقاله را میتوانید از طریق این لینک مشاهده کنید"
به طور کلی، آنتی بادی ها دیگر برای محصولات پخته شده جالب و مورد توجه نیستند. همچنین، آزمایش های مبتنی بر کشت می توانند زمانبر، نادقیق و نیازمند نیروی کار زیاد باشند. پس از توسعه واکنش زنجیره ای پلیمراز (PCR) و به ویژه Real-time-PCR) RT-PCR)، این تکنیک ها به دلیل سرعت و حساسیت بالا در شناسایی گونه ها به عنوان روش آنالیز مواد غذایی انتخاب شدند.
«واکنش زنجیرهای پلیمراز» یک تکنیک رایج آزمایشگاهی است که در تکثیر قطعه مشخصی از DNA به کار میرود. هیچ محدودیتی در انتخاب قطعه مورد نظر وجود ندارد و ممکن است حاوی تمام یا بخشی از ژنهای رمزکننده پروتئين، RNA یا بخشی از یک بیومارکر باشد. در روش RT-PCR امکان رویت لحظه به لحظه یک واکنش مهیا است و در هر دوره ( سیکل) ، کاربر میتواند پروسه تکثیر را بررسی کند.
با این حال، یکی از نقاط ضعف عمده PCR، حساسیت DNA است که میتواند طی فراوری و تولید مواد غذایی تخریب شود. به عنوان مثال، حرارت دادن ملایم نشان داد که تکنیکهای مبتنی بر PCR برای محصولات غذایی پخته یا فراوری شده محدودیت دارند، به ویژه اگر نمونه به اندازه کافی کوچک نباشد. روشهای پروتئومیک مبتنی بر اسپکترومتری جرمی (MS) به طور فزایندهای برای تشخیص اختصاصی مقادیر ناچیز یک ماده در محصولات غذایی استفاده میشوند. به طور کلی، پروتئینها یا پپتیدهای خاصی در روشهای MS شناسایی میشوند و در برابر فراوری کاملاً پایدار هستند اما ممکن است نیاز به جداسازی پیچیده مبتنی بر ژل پروتئومهای غذایی داشته باشند.
تمایل فزاینده محیط زیستی برای جایگزین کردن پروتئینهای حیوانی، بخش خوراک دام و مواد غذایی اروپا را به سمت منابع پروتئینی نوآورانهای از جمله حشرات سوق داده است. امروزه منابع پروتئین جایگزین به شدت مورد نیاز هستند زیرا مساحت زمین در دسترس برای برآورده کردن تقاضای رو به رشد گوشت کافی نیست. این در حالیست که تولید فعلی گوشت در حال حاضر تأثیر قابل توجهی بر تغییرات آب و هوایی دارد و مسئول حدود 10٪ گازهای گلخانهای تولید شده در اروپا است. با توجه به مزایای کیفی زیست محیطی و در صورت مصرف و مدیریت درست، حشرات خوراکی برای مصرف انسان و حیوان ایمن در نظر گرفته میشوند و در صورت رعایت سایر توصیه های غذایی رایج، برای یک رژیم های غذایی با کیفیت بسیار مفید هستند.
با این حال، همانند سایر مواد غذایی خطرات میکروبی، شیمیایی، فیزیکی، آلرژیزایی، انگلی و سمشناسی هنگام انتخاب حشرات برای مصرف انسان یا حیوان، باید مد نظر قرار گیرند. نیاز به تحقیقات جزئیتر در مورد مسائل ایمنی غذا و خوراک مرتبط با حشرات خوراکی وجود دارد تا از بروز خطرات جلوگیری شود و تدوین قوانین سازنده برای تنظیم استفاده از حشرات به عنوان غذا و خوراک را هدایت کند.
در حال حاضر، پروتئینهای فرآوری شده حیوانی از هفت نوع حشره به موجب مقررات کمیسیون اتحادیه اروپا شماره 2017/893 از اول ژانویه 2017 در اتحادیه اروپا مجاز شدهاند و تنها در خوراک آبزیان استفاده میشوند. در مورد مصرف انسانی، همه تولیدکنندگان غذای حشرات باید از اول ژانویه 2018 یک برنامه غذایی نوین که مختص محصولاتشان است و شامل نوع گونه، زمان برداشت، بستر مورد استفاده برای پرورش، روشهای پرورش و فرآوری و غیره را تکمیل کنند. با توجه به استفاده روزافزون از حشرات خوراکی در غذا و خوراک در آینده، شناسایی گونههای حشرات یک کاربرد نوظهور برای کنترل مؤثر تقلب در محصولات حشرات و نقض الزامات برچسبگذاری محصولات حاوی حشرات است.
واقعیت این است که تاکنون صرفا تحقیقاتی در زمینه تشخیص حشرات در ماتریسهای غذایی یا خوراک انجام شده است که بر روی تکنیکهای PCR و یک مطالعه پروتئومیک تمرکز دارند. Tramuta نشان داد که یک مجموعه از PCRهای چندگانه (mPCRs) که بر پایه پرایمرهای موجود در ژن 16s rRNA ایجاد شدهاند، شناسایی سریع ژنتیک گونههای مختلف حشرات را ممکن ساخته و این روش حساس، ساده و افتراقی (تمیزکننده) است.
پرایمر در یک توالی کوتاه نوکلئیک اسید است که یک نقطه شروع برای سنتز DNA فراهم میکند. همچنین ژنی که معتبرترین اطلاعات ژنتیکی را برای بررسی ارتباطات تکاملی در دسترس محققان قرار میدهد، ژن 16S rRNA نام دارد. این ژن توالی از DNA است که بخش RNA زیر واحد کوچکتر ریبوزوم باکتری را رمزگذاری میکند.
این مطالعه بر روی 9 گونه حشره، از جمله سه گونه امیدوارکننده برای تولید صنعتی در غذای انسان، یعنی mealworm ، lesser mealworm و یک گونه جیرجیرک از جنس Gryllus L. 1758 متمرکز شد. Debode دو ژن وینگلس و کادرین را برای شناسایی کمیتی Tenebrio molitor در ماتریس غذایی پیشنهاد کرد، در حالی که مارین یک تشخیص بر پایه PCR روی ژن COX3 برای Hermetia illucens L. 1758 که یک گونه حشره امیدوارکننده در تولید پروتئین خوراکی است توسعه داد.
mealworm در واقع لارو نوعی سوسک به نام سوسک آرد است. بعد از اینکه تخم گذاری توسط این سوسک سیاه مولد انجام میگیرد، تخمها در محیطی کاملا تمیز بر روی بستر سبوس گندم و جو تخم ریزی شده و با فراهم کردن شرایط دمایی و رطوبت کافی بعد از چند روز از تخم خارج شده و به صورت شفیره هستند. شفیره mealworm به مرور از همین بستر تغذیه کرده و بزرگ میشود که پس از گذشت کمتر از دو ماه که به مرحلهای میرسد که ظاهری شبیه کرم دارد و به آن mealworm گفته میشود.
این دو مطالعه نتایج مثبت کاذب کمی را در مورد برخی از گونههای دیگر حشرات نشان دادند و با توجه به اینکه تعداد زیادی از حشرات در طبیعت یافت میشوند، خطر نتایج مثبت کاذب در نتیجه واکنش نامطلوب با توالی غیرهدف هنگام استفاده از روش های مبتنی بر PCR قابل حذف نیست.
Belghit نشان داد که شاتگان پروتئومیک در ترکیب با مقایسه طیفی مستقیم قادر است آرد حشرات تجاری را بر اساس طبقهبندی تاکسونومیک آنها تمایز دهد. دستهبندی پایگاه داده طیفی شناسایی پروتئینها و پپتیدهای اختصاصی حشرات برای چهار گونه هدف را ممکن کرد.
هدف نهایی شاتگان پروتئومیک شناسایی پپتیدها و در نتیجه تشخیص و تعیین مقدار پروتئینهای موجود در نمونه است.پیشرفتهای اخیر در زمینه روشهای مختلف جداسازی از جمله معرفی بسترهای جدید و جداسازیهای چند بعدی، طیف سنج جرمی و همچنین طراحی الگوریتمهای نوین جستجو در پایگاههای اطلاعاتی رسیدن به این امر را تسهیل کرده است.
همانطور که Belghit پیشنهاد داد، نیاز به غنیسازی پایگاه دادههای پروتئینها و پپتیدهای حشرات خوراکی وجود دارد و این مطالعه پتانسیل استفاده از آنالیز پروتئومیک LC-MS را برای تشخیص چهار گونه حشره خوراکی در آردهای تجاری حاوی حشرات در دسترس بررسی میکند.
۲- مواد و روشها
۱-۲- آماده سازی حشرات
دو منبع برای هر گونه مدل انتخاب شد: نمونههای خشکشده از آزمایشگاه حشرهشناسی عملکردی و تکاملی Agro-Bio Tech (دانشگاه Liège) و آردهای تجاری به دست آمده از تماسهای مستقیم با تولیدکنندگان یا تامین کنندگان از طریق پلتفرم بینالمللی حشرات برای غذا و خوراک. چهار گونه بر اساس در دسترس بودن در هر دو منبع انتخاب شدند:
T. molitor، A. diaperinus، G. assimilis و H. illucens.
حشرات کامل در آسیاب لوله ایکا با سرعت 15000 دور در دقیقه در مدت 2 دقیقه آسیاب شدند. آردهای حشرات بر اساس توصیه های استاندارد (ISO 24276 (2006 نمونه برداری شدند.
ابتدا 5 گرم آرد حشره تهیه شد و سه نمونه 200 میلیگرمی برای مخلوط کردن با بافر استخراج و انکوبه کردن در یخ به مدت 40 دقیقه انتخاب شدند. مواد سانتریفیوژ شدند و مایع رویی به لوله جدید منتقل شد تا با سیستم تجاری 2D - Clean up خالصسازی شود.
بهطور خلاصه، خالصسازی با اضافه کردن 300 میکرولیتر بافر رسوبدهی و پس از آن انکوباسیون در یخ به مدت 15 دقیقه انجام شد. پس از آن، 300 میکرولیتر ماده رسوبدهنده اضافه و نمونه مجدداً سانتریفیوژ شد.
مایع رویی دور ریخته شد و 40 میکرولیتر از ماده رسوبدهنده روی رسوب برداشته شد. به رسوب اجازه داده شد تا به مدت 5 دقیقه در یخ تهنشین شود. نمونه مجدداً سانتریفیوژ شد و رسوب در 25 میکرولیتر Milli-Q، یک میلی لیتر بافر شستشو و 5 میکرولیتر افزودنی شستشو بازیابی شد.
نهایتا روش خالصسازی با سانتریفیوژ برای بازیافت رسوب پروتئینی انجام شد. میزان پروتئین (بر حسب میکروگرم بر گرم نمونه) برای حشرات کامل و آرد تجاری به شرح زیر بود؛
32.58 ± 1.99 and 15.79 ± 0.65 from T. molitor
56.58 ± 4.25 and 50.19 ± 10.08 from H. illucens
39.81 ± 7.40 and 24.30 ± 5.39 from A. diaperinus
25.45 ± 2.05 and 27.22 ± 4.60 from G. assimilis
نهایتا سه تکرار برای تزریق تکی در آنالیز LC-MS برای هر گونه حشره ادغام شدند.
۲-۲- آنالیز LC-MS
رسوب بازیافتی برای هر نمونه در بیکربنات آمونیوم مجدداً معلق شد و 15 میکروگرم (بر اساس محتوای پروتئین کل تعیین شده با استفاده از کیت کمیسازی RC DC) از آن با 0.2 میکرولیتر دیتیوتریتول (DTT) احیا شد، سپس با 1.47 میکرولیتر یدواستامید 550 میلیمولار آلکیله شد و سپس با تریپسین هضم شد تا نسبت نهایی تریپسین/پروتئین 1/20 (وزنی/وزنی) به دست آید. پس از توقف هضم با اضافه کردن اسید تری فلورواستیک 0.5%، نمونهها تحت خلاء با استفاده از SpeedVac خشک شدند. ترکیب هضم پروتئین مجدداً در آب اسیدی شده با 0.1% TFA معلق و 1 میکروگرم از پپتیدهای بازیافت شده به سیستم LC تزریق شدند.
آنالیزهای LC-MS/MS روی دستگاه Acquity M-Class UPLC متصل به Q Exactive Plus Thermo Scientific، در حالت یونیزاسیون الکترواسپری مثبت انجام شد. ستون تله Symmetry C18 5 μm و ستون تجزیه HSS T3 C18 1.8 μm بود.
نمونهها با سرعت 20 میکرولیتر/دقیقه روی ستون تله در 98% حلال A به مدت 3 دقیقه بارگذاری و سپس روی ستون تجزیه با سرعت جریان 600 نانولیتر/دقیقه با گرادیان خطی زیر جداسازی شدند: شرایط اولیه 98% A؛ 5 دقیقه 93% A؛ 30 دقیقه 60% A؛ 33 دقیقه 15% A که در آن حلال A شامل 0.1% اسید فرمیک در آب و حلال B شامل 0.1% اسید فرمیک در استونیتریل است. مدت زمان کل اجرا 60 دقیقه بود.
روش اسپکتروسکوپی جرمی یک روش TopN-MS/MS است که در آن N روی 12 تنظیم شده بود، به این معنی که اسپکترومتر یک طیف MS کامل را ثبت میکند و 12 پیک شدید در این طیف را انتخاب کرده و یک طیف MS2 کامل از هر 12 ترکیب ثبت میکند.
پارامترهای ثبت طیف MS به شرح زیر است:
دامنه جرمی از 400 تا 1750
قدرت تفکیک 70000
زمان تزریق بیشینه 50 میلیثانیه
پارامترهای ثبت طیف MS2 به شرح زیر است:
پنجره جداسازی 2.0
انرژی برخورد نرمالشده 25
قدرت تفکیک 17500
زمان تزریق بیشینه 50 میلیثانیه
دادههای بدست امده در پایگاه های داده جستجو شدند. پارامترهای اصلی جستجو عبارت بودند از:
حد مجاز 5 ppm برای جرم پیشماده و 20 mmu برای اجزای MS/MS، آنزیم: تریپسین (اختصاصی)، حداکثر 2 شکاف از دست رفته مجاز، اکسیداسیون متیونین به عنوان تغییر متغیر و کربامیدومتیلاسیون سیستئین به عنوان تغییر ثابت.
۳-۲- آنالیز پروتئین
برای ساخت شکلهای ۱ و ۲، هر پروتئین شناسایی شده به صورت انفرادی در نظر گرفته شد تا تنوع کلی از جمله تنوع موجودات مرتبط نیز ارزیابی شود. تعداد پروتئینها در شکل ۱ (۳۶۸ عدد) به کل اختصاصهای بهدستآمده از حشرات مربوط میشود. از آنجایی که برخی از آنها با کدهای دسترسی متفاوت به یک پروتئین اختصاص یافته بودند، اختصاصهای مشابه به یک پروتئین اجماع (ادغام) شدند. این کار تعداد اختصاص ها را برای ۴ گونه حشره به ۲۳۴ در شکل ۲ کاهش داد.
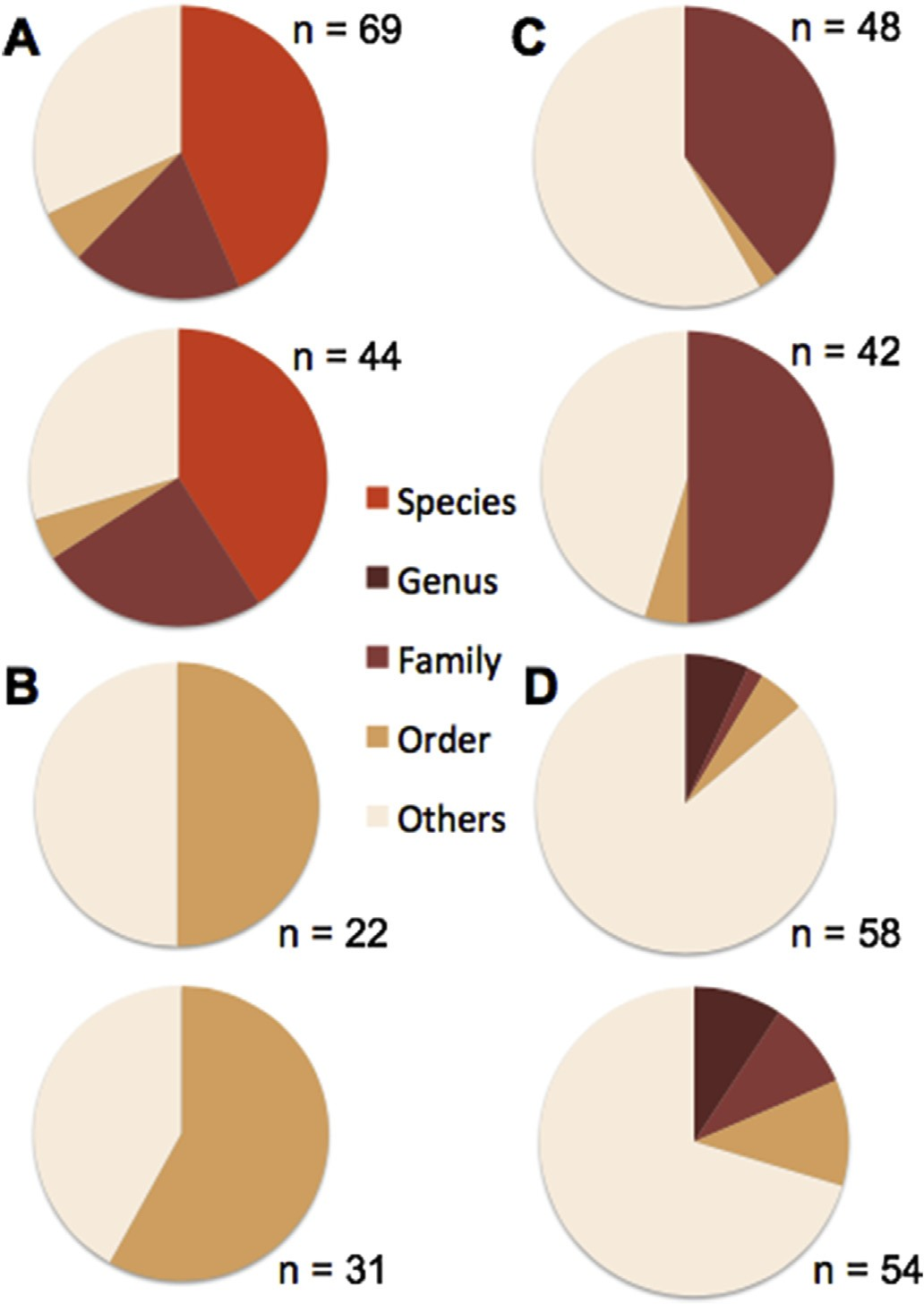
شکل ۱) درصد موفقیت شناسایی پروتئین در نمونه های حشرات بر اساس نزدیکی تاکسونومیکی با گونه حشره مرتبط با پروتئین در پایگاه داده.
Tenebrio molitor (A)، Hermetia illucens (B)، Alphitobius diaperinus (C) و Gryllus assimilis (D)
برای هر گونه حشره، ارقام اول و دوم به ترتیب مربوط به پودر حشرات کامل و آرد تجاری بودند.
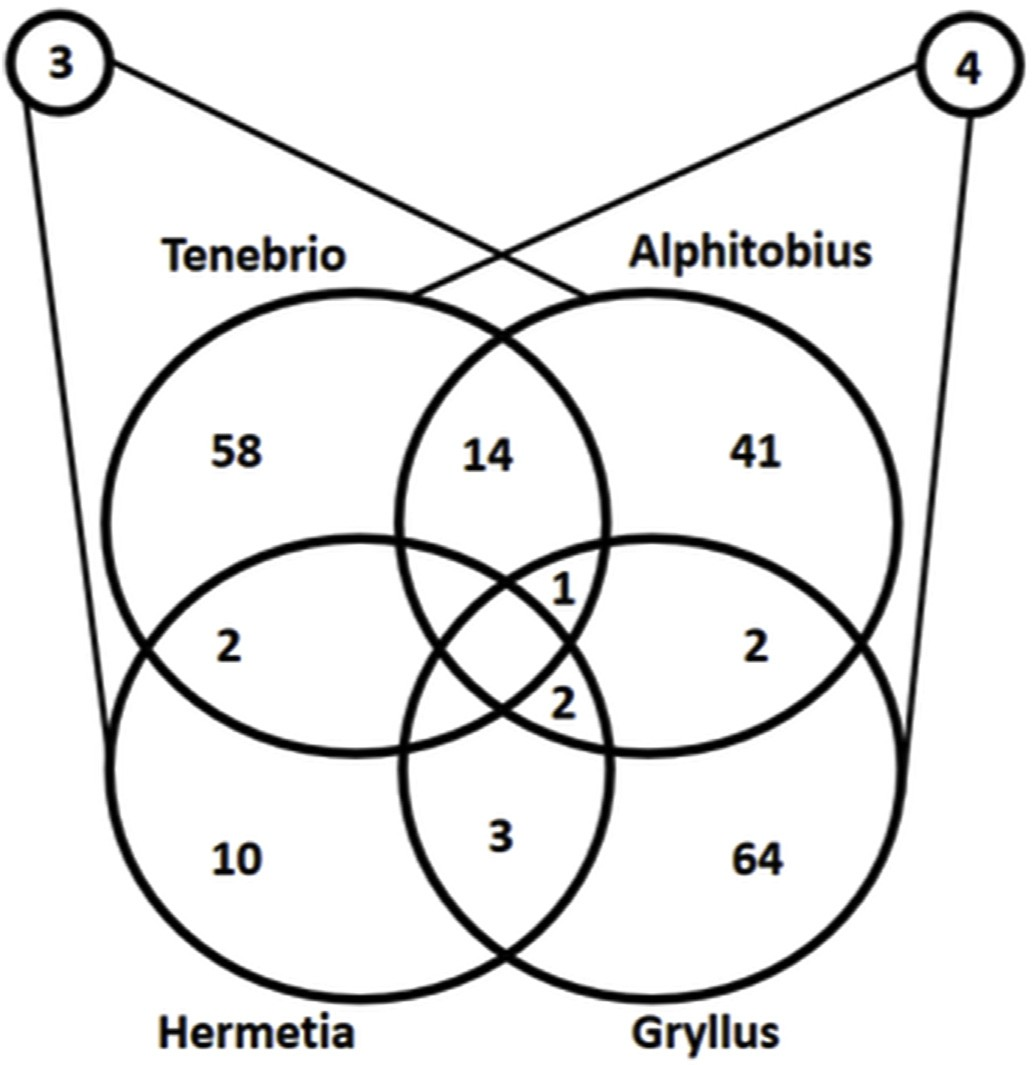
شکل ۲) نمودار ون مقایسه بین شناسایی پروتئینهای مرتبط با هر گونه حشره خوراکی، یعنی Tenebrio molitor، Alphitobius diaperinus، Gryllus assimillis و Hermetia illucens را نشان میدهد.
از آنجایی که فعل و انفعالات بین T. molitor، G. assimilis و بین H. illucens و A. diaperinus نیاز به نمایش سه بعدی دارد، مقایسه از نظر پپتیدهای مشترک در بخش بالایی شکل نشان داده شده است.
برای تمرکز روی گروههای پروتئینی خاصی که به دنبالشان بودیم، تصمیم گرفتیم در شکل ۳ توالیها را بر اساس عملکرد پروتئین اصلی (70) با استفاده از تعداد کل پپتیدهای منحصربفرد شناساییشده از آنالیز طیفسنجی جرمی به عنوان پارامتر مطمئن و حساس دستهبندی کنیم.


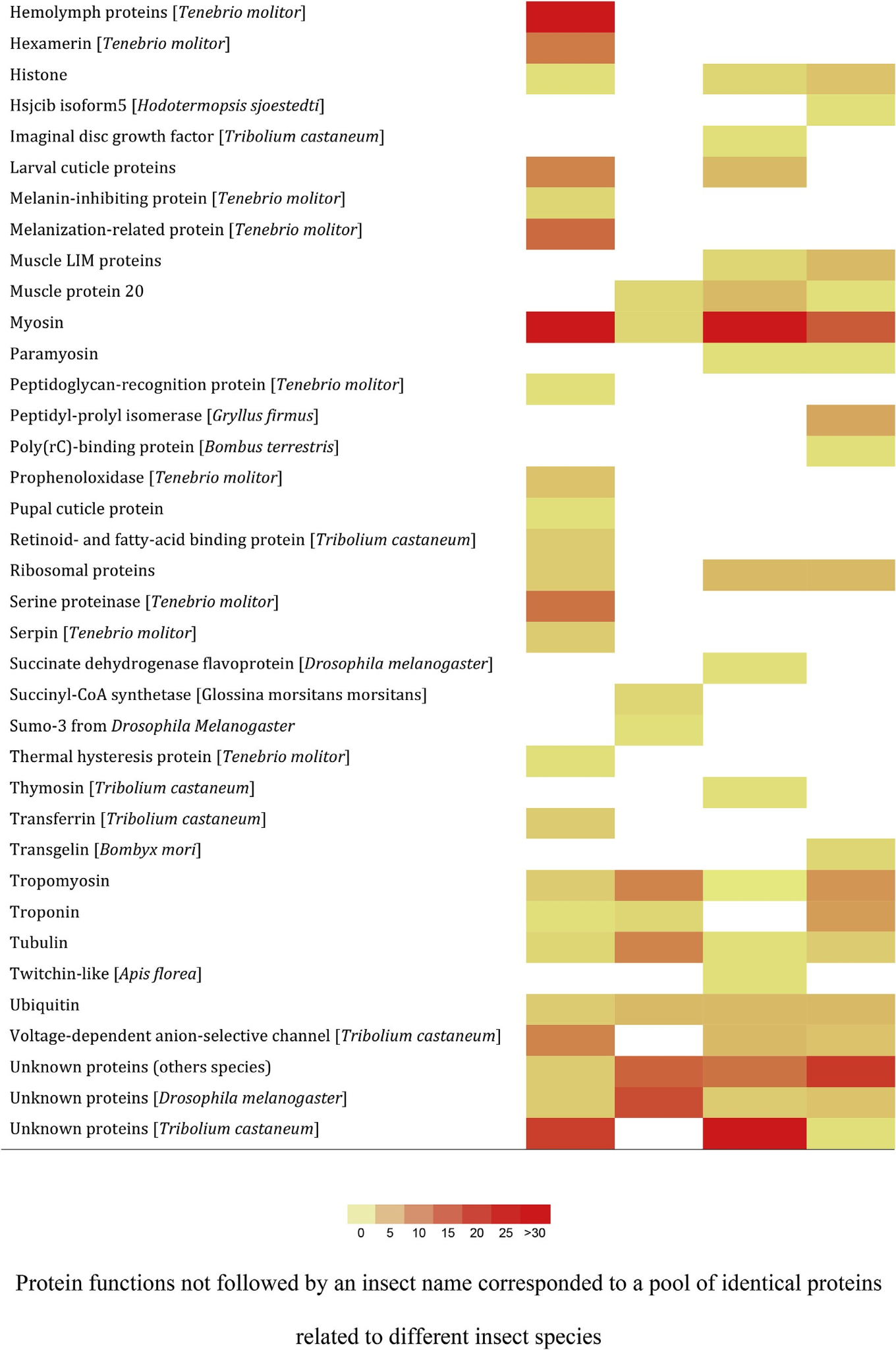
۳- بحث و نتیجه گیری
کارایی شناسایی حشرات در سطح گونه بر اساس دادههای توالی پپتید برای گونههای مختلف، متفاوت است. برای مثال در حالی که T. molitor به راحتی با 30 (حشرات آسیاب شده) و 18 (نمونههای صنعتی) پروتئین اختصاصی قابل شناسایی بود، وضعیت برای H. illucens کاملاً متفاوت بود و در واقع، هیچ شباهتی به آرایه (تاکسون) های نزدیک حشرات در زیر سطح راسته پیدا نشد و نزدیک ترین دوبالان انواعی از مگسها بودند.
در میان این دو مورد افراطی (T. molitor با تعداد زیاد پروتئینهای قابل شناسایی و H. illucens بدون هیچ شباهتی)، برای A. diaperinus، پپتیدها به 19 پروتئین در نمونه حشرات آسیاب شده و 21 پروتئین در نمونه تجاری شباهت نشان دادند که مربوط به موجودات خانواده Tenebrionidae بودند.
در نهایت، برای G. assimilis اکثر شباهتهای مشاهده شده با تاکسونهای دور از نظر خویشاوندی بود، اما چند پروتئین (4 تا در نمونه حشرات آسیاب شده و 5 تا در نمونه تجاری) به سطح سرده Gryllus نزدیکتر بودند.
تنوع کارایی مرتبط با چهار گونه حشره آزمایششده به طور مستقیم با وضعیت هر حشره خوراکی هدف از نظر میزان توجه مولکولی کلی و به خصوص دسترسی به ژنوم توالیشده مرتبط است.
به طور کلی شناسایی پروتئین با اندازه پایگاه دادههای پروتئوم مرتبط با هر گونه وابسته است و توسط آن محدود میشود.
بسیاری از پروتئینهای شناسایی شده در این پژوهش با ژنوم توالیشده Mealworm مرتبط بودند، به خصوص برای نمونههای A. diaperinus و T. molitor. همچنین چندین شناسایی دیگر هم با یکی از اولین ژنومهای در دسترس یعنی ژنوم نوعی مگس که به پروتئینهای H. illucens شبیه بود، صورت گرفت. در رویکرد مبتنی بر PCR، هیچ پروتئین خاصی برای H. illucens و سایر کاربردهای پروتئومیک مرتبط با این حشره برای شناسایی آن پیشنهاد نمیشود. به طور کلی ما قادر به ارائه راه حل مؤثری برای این مدل گسترده که برای خوراک طیور و آبزیان توسعه یافته است، نیستیم.
در آنالیز LC-MS، زمانی که نتایج حاصل از دو نوع منبع حشره، یعنی نمونههای کامل زمینه و آرد تجاری برای هر یک از چهار گونه حشره آزمایششده مقایسه شدند تکرارپذیری بیشتر به وضوح مشاهده شد.
وقتی پروتئینهای شناسایی شده برای 4 گونه حشره هدف با هم مقایسه شدند، همپوشانی (اشتراک) بسیار کمی بین آنها مشاهده شد. از مجموع 234 پروتئین شناسایی شده، تنها 21 پروتئین مشترک بین حداکثر 2 گونه وجود داشت و تنها 3 پروتئین بین 3 گونه به اشتراک گذاشته شده بود. یعنی تقریبا هیچ همپوشانی پروتئینی بین 4 گونه حشره وجود نداشت. همپوشانی کم پروتئینی مشاهده شده بین 4 گونه حشره، امکان شناسایی پروتئینهای خاص و منحصربفرد برای دو گونه سوسک (T. molitor و A. diaperinus) و یک گونه ملخ (G. assimilis) را که در این مطالعه بررسی شده بودند، فراهم کرد.
چند پروتئین خاص و منحصر به G. assimilis در پایگاه داده NCBI شناسایی شدند.
۱- یک پروتئین غده فرعی با ۱۰ پپتید
۲- یک پپتیدیل-پرولیل ایزومراز با ۳ پپتید
۳- یک پروتئین شوک حرارتی ۹۰
۴- یک پپتیدیل-پرولیل ایزومراز با ۲ پپتید
پروتئین شوک حرارتی ۹۰ یا HSP90 یک پروتئین محافظتکننده است که در شرایط استرسی در سلول بیان میگردند. نقش این سلولها جلوگیری از تغییر کونفورماسیون پروتئینها تحت عوامل استرسی میباشند. همچنین پروتئین غده فرعی گروهی از پروتئینها هستند که توسط غدد فرعی موجود در دستگاه تولیدمثلی نر برخی حشرات ترشح شده و نقش مهمی در فرایند جفتگیری و باروری دارند. پپتیدیل-پرولیل ایزومراز آنزیمی است با تسهیل فرآیند تاخوردگی و شکلگیری ساختار سوم پروتئینها، نقش مهمی در تنظیم بیان ژن و همچنین ترمیم و بازسازی پروتئینهای آسیب دیده دارد.
با توجه به اینکه هنوز اطلاعات جامع و کاملی از ژنوم حشرات در دسترس نیست، شناسایی و تمرکز روی پروتئینهای فراگیر و مشترک در حشرات میتواند راهبرد مناسبی برای تشخیص حضور آنها در مواد غذایی و خوراک باشد. چون این پروتئینها در اکثر حشرات وجود دارند میتوان با شناسایی آنها، حضور حشرات را تا حدودی تشخیص داد.
با توجه به اینکه هنوز اطلاعات جامع و کاملی از ژنوم حشرات در دسترس نیست، شناسایی و تمرکز روی پروتئینهای فراگیر و مشترک در حشرات میتواند راهبرد مناسبی برای تشخیص حضور آنها در مواد غذایی و خوراک باشد. چون این پروتئینها در اکثر حشرات وجود دارند میتوان با شناسایی آنها، حضور حشرات را تا حدودی تشخیص داد.
در مرحله فعلی، شناسایی پروتئینهای اختصاص یافته به هر حشره، نمیتواند به طور کامل و دقیق روشی برای شناسایی گونه آن حشره باشد. چون پایگاهدادههای موجود هنوز برای شناسایی قطعی و مطلق گونههای حشرات، ناقص و ناکامل هستند. یعنی اطلاعات لازم درباره همه گونههای حشرات و پروتئینهای اختصاصی آنها در دسترس نیست و نیاز به تکمیل و گسترش پایگاههای داده وجود دارد.
همچنین چند پروتئین به صورت اختصاصی به T. molitor نسبت داده شده بود که میتوانند به عنوان اهداف جالبی برای شناسایی این گونه مد نظر قرار بگیرند:
۱- پروتئین القاکننده کپسوله شدن با ۱۹ پپتید
۲- پروتئین مرتبط با ملانیزاسیون با ۱۵ پپتید
۳- هگزامرین با ۱۳ پپتید
۴- پروفنل اکسیداز با ۱۲ پپتید
۵- پروتئین همولنف با ۹ پپتید
همچنین دو پروتئین آلفا آمیلاز و کاتپسین L مانند (با ۷ پپتید) هم قابل شناسایی هستند.
پروتئین های القا کننده کپسوله شدن و مرتبط با ملانیزاسیون پروتئین هایی هستند که در واکنش های دفاعی حشرات نقش دارند.
هگزامرینها پروتئینهای ذخیرهای حشرات بوده و منبعی از اسیدهای آمینه هستند که طی دوران لاروی و رشد، مصرف میشوند تا لارو به حشره بالغ و کامل تبدیل گردد.
پروفنل اکسیداز یک آنزیم در سیستم ایمنی حشرات است و زمانی که یک حشره دچار عفونت میکروبی میشود، این آنزیم فعال میشود.
چند پروتئین که مربوط به T. castaneum هستند، برای شناسایی گونه A. diaperinus نیز مفید بودند. این پروتئینها عبارتند از:
۱- پروتئین TcasGA2 با ۱۱ پپتید (یک پروتئین همولنف هست که نقش مهمی در متابولیسم قندها در حشرات دارد)
۲- ATP سنتاز با ۸ پپتید (آنزیمی که ADP را به ATP تبدیل میکند.)
۳- آلفا آکتینین و یک پروتئین شوک حرارتی با ۶ پپتید
۴- پروتئین کوتیکولی
۵- کاتالاز با ۵ پپتید
۵- پروتئین LIM عضلانی با ۴ پپتید
جنس بدن حشرات از کوتیکول است که مادهای کیتینی است و بخش غیر زنده پوست حشرات را تشکیل میدهد. اگزوکوتیکول از کیتین و آرتروپودین (پروتئین مخصوص بندپایان) تشکیل شدهاست.
پروتئین LIM عضلانی یک پروتئین ساختاری مهم در سلولهای عضلانی است. پروتئین LIM عضلانی با اتصال به پروتئینهای سازهای دیگر، نقش بسیار مهمی در الگودهی، تکامل و تمایز سلولهای عضلانی و نیز تنظیم انقباض آنها دارد.
به طور خلاصه، استفاده از روش پروتئومیکس مبتنی بر کروماتوگرافی مایع-طیفسنجی جرمی (LC-MS) را میتوان به عنوان یک روش بالقوه و مفید برای بررسی ترکیبات و اجزای غذاها از نظر وجود یا عدم وجود حشرات خوراکی در آنها معرفی کرد. این روش میتواند با شناسایی پروتئینها و پپتیدهای اختصاصی، وجود انواع حشرات در یک ماده غذایی را تایید یا رد کند.
برای توسعه روتین و معمول این راهبرد، هر دو رویکرد زیر باید بیشتر مورد بررسی قرار گیرند:
۱) رویکردهای مستقل از پایگاه داده، همانطور که در مطالعه Belghit و همکاران (2019) ارائه شده است.
۲) پپتیدهای انتخاب شده به عنوان نشانگر، همانطور که در اینجا نشان داده شده است.
هدف از بررسی بیشتر این دو رویکرد، استفاده از آنها برای تشخیص حضور یا آلودگی ناشی از حشرات در نمونهها است.
گونههای سوسک مانند Mealworm را میتوان با دانش فعلی از پایگاه داده پپتیدی، به سرعت بررسی و شناسایی کرد. برای رسیدن به دقت بیشتر در شناسایی گونههای ملخ با استفاده از آنالیز پروتئینی، نیاز است تا آزمایشهای بیشتری روی چند گونه از جنسهای Gryllus و Acheta انجام شود و نتایج مقایسه گردد. همچنین شناسایی دقیق گونه H. illucens نیز تنها پس از تعیین توالی ژنوم آن امکانپذیر خواهد بود؛ چرا که اکنون اطلاعات ژنتیکی و پروتئینی اندکی در مورد این گونه و به طور کلی در مورد خانواده Stratiomyidae موجود است.
همچنین باید به مکمل بودن احتمالی رویکردهای پروتئینی و DNAای (ژنومی) اشاره کنیم. پروتئینهایی که برای گونههای مختلف حشرات توسط اسپکترومتری جرمی شناسایی میشوند، میتوانند به طور اولویتدار در سطح توالی DNA اصلی خود مورد بررسی قرار گیرند تا مشخص شود که آیا میتوانند به عنوان حمایتی جالب برای توسعه یک آزمون PCR در زمان واقعی اختصاصی برای حشره مورد نظر باشند یا خیر.
یعنی ابتدا پروتئینهای خاص هر حشره با روش طیفسنجی جرمی شناسایی میشوند، سپس توالی DNA متناظر آن پروتئینها بررسی میشود تا مشخص شود آیا میتوان از آنها به عنوان هدف برای آزمون PCR در زمان واقعی جهت تشخیص آن حشره استفاده کرد یا خیر.




